Plant Cell Energy Transductions 3
Biological Oxidations:
Respiration is a process of biological oxidation where various reserve food materials like starch, fats, proteins and their organic compounds are enzymatically converted to simpler products which are ultimately oxidized to CO2 and H2O. During oxidative steps, the energy present in the chemical bonds in the above said components is transferred in the form of high energy bonds of rATP, rGTP and other energy rich molecules. There are other energy rich molecules such as NADH2/NADPH2; they have three time the energy found in ATP and GTPs. Majority of the organisms living on this planet use ATP as the main source of chemical energy, perhaps the only source.
But compounds like GTP, UTP, PEP, CTP, DIPGA, creatine phosphate, etc., are also used by specific energy requiring metabolic pathways or signaling pathways. Some of the compounds require the inputs of the following energy rich compounds

https://the dairyof5biochemains.wordpress.com/2014

Metabolic+pathways+of+carbohydrates; image; cannot understand; http://pixcooler.com

Life at the cellular level can be defined as a network of integrated and carefully regulated metabolic pathways, each contributing to the sum of activities that a cell must carry out. Major Metabolic Pathways; http://reasonandscience.heavenforum.org/

Carbohydrate Metabolism; Cellular process in biosynthesis (Anabolism and degradation (Catabolism); a GENERAL VEIW; http://www.lookfordiagnosis.com/

Different forms of energy rich compounds; www.dwb.unl.edu; http://www1.biologie.uni-hamburg.de/

One of the most energy rich compound used in most of the pathways is ATPs, its beta and gamma are energy rich bonds; www.nature.com


Beta and Gamma P-O-P sigma bonds hold 7.3kcal per bond each or 30.5 kjoules each per mole; www.chemwiki.ucdavis.edu

Inputs for ATP; http://www1.biologie.uni-hamburg.de/

This is the two-dimensional (ChemDraw) structure of ATP (Adenosine Triphosphate). The removal of one phosphate group (green) from ATP requires the breaking of a bond (blue) and results in a 7.3k.cal/mole of release of free energy. Removal of this phosphate group (green) results in ADP, adenosine diphosphate.
Glucose is a basic six carbon monosaccharide which contains trapped solar energy in the form of chemical energy, call it as chemical bonds. All living organisms require chemical energy for various biological activities and the same is supplied by the oxidation of glucose or similar carbon compounds including lipids. Thus, glucose is at the hub of energy supplying metabolic pathways. The stepwise oxidation of glucose not only yields energy rich molecules but also provides many intermediary carbon skeletons for the synthesis of other components, like amino acids, nitrogen bases, nucleotides, lipids, etc. Based on the utilization or non-utilization of oxygen, respiratory process is classified into aerobic type and anaerobic type.

The mechanism of biological oxidative process varies and it depends upon the organism involved, the metabolic status of the said organism and the availability of oxygen in the immediate environment. In the presence of oxygen, most of the eukaryotic organisms respire aerobically, but in the absence of it, some like yeast cells resort to anaerobic type of respiration for their survival. But some organisms are obligate anaerobes. The anaerobic respiration results in the formation of ethanol and in some cases, lactate is produced. However the glycolytic pathway of biological oxidation is common for both aerobic and anaerobic processes. In aerobic reactions, the glycolytic products such as pyruvates are subjected to Kreb’s cycle events and terminal oxidation. On the other hand, in anaerobic respiration, the glycolytic products are converted to either ethanol or lactate which depends upon the organism involved. A part of the oxidation process takes place in cytoplasm, and the other steps in Mitochondria.

http://classes.midlandstech.edu/


http://www.desktopclaasses.com; http://ilgiliforum.com/

Damien Arnoult, Fraser Soares, Ivan Tattoli, Stephen E Girardin; http://embor.embopress.org/

Innate immune signaling at mitochondria; http://embor.embopress.org/

Mitochondrial release Cyt C leads to cell apoptosis; http://embor.embopress.org/

Hypothetical models of the role of contacts between mitochondria and ER in apoptosis; The hFis1/Bap31 platform transmits the mitochondrial stress signal to the ER via the activation of procaspase-8. The cytosolic region of the ER integral membrane protein Bap31 is cleaved by activated caspase-8 to generate proapoptotic p20Bap31, which causes rapid transmission of ER calcium signals to the mitochondria via the IP3 receptor. At close ER-mitochondria contact sites, mitochondria take up calcium into the matrix via the mitochondrial calcium channels MICU1 or LETM1. The massive influx of calcium leads to mitochondrial fission, cristae remodeling, and cytochrome release. Mfn2 is enriched in the mitochondria-associated membranes (MAM) of the endoplasmic reticulum (ER), where it interacts with Mfn1 and Mfn2 on the mitochondria to form inter-organellar bridges. Upon apoptosis signal, a BH3-only member of the Bcl-2 family, Bik, induces Ca2+ release from the ER and, in turn, induces Drp1 recruitment to the mitochondria and their fragmentation and cristae remodeling. SERCA, sarco-/endoplasmic reticulum Ca2+-ATPase. MICU1, mitochondrial calcium uptake 1. LETM1, leucine zipper/EF hand-containing transmembrane 1.; http://www.hindawi.com/

Mitochondrial handling; http://physiologyonline.physiology.org/;
Ca2+ cycling between the extramitochondrial (cytosolic) and mitochondrial martrix space. Import and export of Ca2+ is coupled to H+ and Na+ cycling maintained by electron transport chain in mitochondria.

Summary of substances targeting ER, Mitochondria, Oxidative stress and altered Ca2+ homeostasis.; http://www.frontiersin.org/

ER mitochondria Ca2+ cycle (EMRMCC); http://www.frontiersin.org/
AEROBIC RESPIRATION
The most important energy source for plants is starch and for animal’s glycogen or fat, again it depends upon the stage and status of the organ and organism. But in plants during germination, seeds utilize starch or fatty acids they store. To start with, the complex compounds are hydrolyzed to simple forms like glucose or fructose, acetyl CoA, or derivative of amino acids, which are drawn into catalytic pathway that operates in cytosol later drawn into mitochondria. All the required enzymes are found in cytosol.

The formation of hexose phosphates by hexokinase is therefore a very important reaction in the metabolism of carbohydrates. Transgenic studies with a cytoplasmic hexokinase in potatoes by Veramendi et al. (1999), provides useful insight on in vivo sugar metabolism. They were able to achieve large increases and decreases in hexokinase activity in leaves and tubers using sense and antisense constructs. This did not affect tuber growth, starch accumulation or other metabolism of tubers. However antisense suppression of hexokinase in leaves resulted in a 3- fold increase in starch, a 2-fold increase in glucose and a decrease in sucrose after a dark period. It has long been thought that export of reduced carbohydrate from chloroplasts was only in the form of triose phosphates. While this may be true in the light, the export of assimilate from transient starch turnover in chloroplasts in the dark appears to be predominately in the form of glucose as indicated in these studies; http://www.uky.edu/

Amount of energy held in each of the covalent bonds; www. Users.rcn.com.
If glucose is the starting material, it stores considerable amount of chemical energy i.e bond energy (during photosynthesis) and the same is released in step wise manner and nearly 30 percent of it is captured and stored in ATP chemical bonds and the rest is released as heat. Glucose is not just used for generation of metabolic energy, but also used for producing various compounds. The process of biological oxidation is same in all organisms irrespective of their kinds and forms.

Average Bond Energies, Kcal/mole (1mole=6.2 x 10^23).
C-H = 98,
O-H = 110,
C-C = 80,
C-O = 78,
H-H = 103,
C-N = 65,
O=O = 116,
C=C = 145
C=O =187,
Glycolysis
This process operates in the cytoplasm but outside the mitochondria. The enzymes needed for this process are found in cytosol and some of them are associated with the outer surface of the outer membrane of the mitochondria as granular particulates.
The glycolytic process takes place in two phases, i.e., first from glucose to phosphoglyceraldehyde and the second from PGALD to pyruvate. These reactions are very well regulated by the end products or by the energy status of the cells.
To begin with, free glucose is raised to higher state of energy by a phosphorylation reaction with the help of an enzyme called hexose kinase, which utilizes a molecule of ATP. This reaction is referred to as priming reaction.
If starch is utilized, the breakdown product of starch-by-starch phosphorylase enzyme is glucose 1-P, which inturn is converted to glucose 6-P by phosphoglucomutase. This enzyme is a regulatory enzyme.
Starch + P1 Phosphorylase Glucose 1-(P)
Glucose 1-(P) glucomutase Glucose 6-(P)
Other hexoses like galactose, mannose, etc., if available in the cells, are first converted to glucose 6-(P) before they are drawn into glycolytic pathway.
Mannose -> Glucose 6–P
Galactose -> Glucose 6-P

www.studyblue.com
Glucose 6-P is then converted to fructose 6-P by phosphoglucose-isomerase. Immediately, the fructose 6-P is further primed up to fructose 1-6 diphosphate by an enzyme called phospho-fructokinase. In this step another molecule of ATP is utilized. This enzyme is allosteric in nature and the cellular ATP level regulates its activity. If the cellular energy level is high i.e. high content of ATP, the enzyme activity is inhibited. On the contrary, if the concentration of ATP is low (i.e. more of ADP) the enzyme is active and proceeds in the formation of fructose 1-6 triphosphate; it is the peak of energy rich product in Glycolysis.

Fructose6P to fructorse1,6-diP is regulated, and it depends upon the availability of ATP. And regulated by active PFK (phospho-fructose kinase) and phosphorylates the F6P to F1,6 diP, otherwise PFK remains inactive, whose activity in turn is controlled by specific kinases.

The priming process is like expending the energy initially to push a big rock on the summit of a mountain to such a position, when it is dropped; it just rolls down all along the slop releasing its energy at every step of its bounced movements. Similarly, further glycolytic reactions of fructose 1-6 diphosphate is a downhill process. Then fructose 1-6 diphosphate is cleaved into two 3 carbon compounds, called glyceraldehyde 3-phosphate (PGALD) and di hydroxy acetone phosphate (DHAP). The enzyme that performs this reaction is fructose diphosphate aldolase which can reverse this reaction by aldol condensation.
Glyceraldehyde 3 phosphate and di hydroxy acetone phosphates are isomers and they can be easily inter-converted by an enzyme named triose phosphate isomerase.
Further reactions depend upon the energy status of the cell. If cells require more energy, DHAP is first converted to PGALD which is then subjected to the second phase of glycolytic reactions. On the contrary, if the cellular ATP molecules are found in sufficient quantities, DHAPs are channeled towards glycerol and lipid production. This part of the reaction is regulated by the ATP and ADP levels of the cells or ATP/ADP ratios.
If more energy is required, the second phase of glycolytic process proceeds where glyceraldehyde 3 phosphate is oxidized to 1.3 di phosphoglycerate. The enzyme glyceraldehyde phosphate dehydrogenase which brings about this reaction utilizes inorganic phosphates and at the same time, its coenzyme NAD gets reduced to energy rich NADH+H. So a part of energy is stored in NADH+H (the energy held in NADH+H is substantial).
PGALD + Pi + NAD ->1-3 PGA (DiPGA) +NADH2
DiPGA + ADP-> ATP + 3’PGA,
3” PGA -> 2’PGA,
2’PGA-> H2O + PEP
PEP + ADP -> Pyruvate + ATP
The DIPGA is an energy rich compound and the same is then subjected to substrate phosphorylation by an enzyme called phosphoglycerate kinase. In this process, the energy rich anhydride phosphate bound is transferred to ADP to produce energy rich ATP molecule. Note the ATP generated is by substrate phosphorylation. The resultant product of this molecule is 3’phosphoglyceric acid, which is then subjected to transfer of phosphate group from 3’ position to 2’ position by phosphoglyceromutase. The product is 2’ phosphoglycerate which is then acted upon by enolase enzyme, where a molecule of water is removed from 2’ carbon position to yield another energy rich compound called phosphoenol pyruvate.
The high energy phosphate bond of phosphoenol pyruvate is then transferred to ADP to produce an ATP by an enzyme called pyruvate kinase. This reaction also produces the ultimate product of glycolytic reaction called pyruvate.
At the end of glycolytic pathway, one mole of glucose yields 2 moles of pyruvates; 2 moles of NADH+H and 4 moles of ATP. Initially 2 moles of ATPs are used for priming reactions of the first phase of glycolytic reactions; the net gain of ATP in these glycolytic reactions is 2 moles of ATP for every mole of glucose.
Glucose + 2 ATP + 2 NAD + 4 ADP + 2PI -> 2 Pyruvate
2 NADH + H + 4 ATP + 2 ADP
Pyruvate a 3 carbon kenotic acid produced at the end of glycolytic pathway is transported from cytosol into mitochondrial matrix, across the mitochondrial membrane.
The fate of the pyruvate depends upon the availability of O2. In aerobic process the pyruvate is drawn to Kreb’s cycle. Instead, if oxygen is not available, certain organisms like yeasts, and lactate bacteria use pyruvate and produce ethanol and lactate respectively by intramolecular biological oxidation.

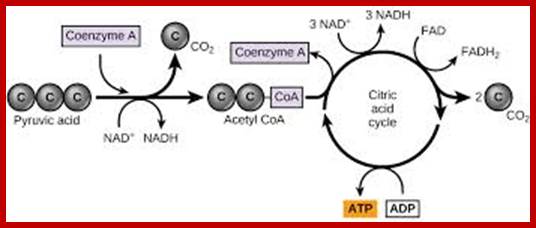
While in transit pyruvate is oxidized to Acetyl-coA by a very important enzyme complex called pyruvate dehydrogenase. Then, 2C acetyl-coA is drawn into a cyclic biochemical pathway called citric acid cycle. As the cyclic reactions were elucidated by Han’s Kreb’s, this pathway is also called Kreb’s cycle.
Entry into Kreb’s cycle or citric acid cycle:
The acetyl Co.A (2C) condenses with another ketonic acid called oxaloacetate (OAA)(4C) with the help of citrate synthase. The product is citric acid, a six-carbon dicarboxylic acid. Citrate synthase is a regulatory enzyme and its activity is regulated by the levels of succinyl Co.A and ATP. In fact, this acts as a pacemaker.


Pyruvate produced at the end of Glycolysis undergoes a series of reactions leading to the production of Acetyl CoA shown below; these reactions are very critical for the entry into Krebs’s cycle. Pyruvate dehydrogenase reactions; www.imgarcade.com; www.peds.ufl.edu

Cut-away model of the fully assembled PDC viewed on its 3-fold axis. The E3 homodimer (red) of A. vineland x-ray structure filtered to 20Å resolution was docked into the pentagonal opening of the core (green) according to studies of the S. cerevisiae PDC. The BP⋅E3 components associated with the core are not revealed in the shaded-surface representation of the bovine kidney PDC at this threshold because only about 6 molecules of E3 are bound. A radial density plot analysis of the complex shows a peak of density inside the core which corresponds to the position of BP⋅E3 peak in S. cerevisiae tE2 The inner linkers (light blue) bind E1 (yellow) to the E2 scaffold (green). The E1-binding site on the E2 inner linker is located ≈50 Å above the scaffold as indicated by “*” and serves as the anchor for the lipoyl domains to pivot. The structure shows that the swinging arm pivots about a position that is ≈50 Å from the E1, E2, and E3 active sites.; http://www.pnas.org/

Production of Acetyl Co A is critical for Citric Acid Cycle or Krebs’s Cycle: http://classroom.sdmesa.edu/

www.voer.edu.vn

The enzyme requires some important vitamins in the form of coenzymes such as NAD, FAD, TPP and Pantothenate. This is a complex of 12 subunits which has been isolated and electro density micrograph is taken.
Pyruvate, a 3-carbon compound is oxidized to two carbon acetyl Co.A. This is a multistep reaction executed by a multiple enzyme complex called pyruvate dehydrogenase.
The Pyruvate dehydrogenase complex is regulated by specific kinases and phosphatases.
There are five steps in which 3 enzymes containing 5 coenzymes take part by dehydrogenation and decarboxylation reactions. Once the substrate, i.e. pyruvate is bound to this multiple enzyme complex, the reaction goes on till the last products produced is released from the surface of the enzyme and some of the intermediate products remain bound to the enzymes.
In the first step, the pyruvate dehydrogenase enzyme removes CO2 from the pyruvate through thiamine pyrophosphate to produce hydroxy ethyl derivate of the thiazole ring of TPP.
E1-TPP+ CH3COCOOH ----> E1-TPP-CH3-CHOH+CO2
In the second step, the hydroxy ethyl group is dehydrogenated resulting in the transfer of acetyl group to the sulphur moiety of lipoic acid, a prosthetic group of enzyme called lipoyl acetyl transferase.
The third step involves enzymatic transfer of acetyl group from the lipoate to thiol group of co-enzyme A. The acetyl Co. A’s formed in this reaction is released from the enzyme surfaces. During this reaction, lipoate reduces dihydrolipoamide.
In the fourth step, lipoamide dehydrogenase removes the hydrogen from dihydrolipoamide, where the co-enzyme FAD gets reduced to FADH2 and lipoid acid is regenerated.
In the fifth step, the hydrogen from the E3 FADH2 (still bound to the enzyme) is transferred to NAD to form NADH+H. Thus, one molecule of pyruvate on dehydrogenation decarboxylation reactions produces one molecule of acetyl Co. A, one molecule of Co2 and one molecule of NADH2. The enzyme pyruvate dehydrogenase, brings about this multistep reaction is a multiple enzyme complex and it is regulated by Ca2+ and ATP. These reactions take place while the pyruvate is transferred across the inner mitochondrial membrane. The carrier for pyruvate is now known to be carnitine phosphate.

www.sphweb.bumc.bu.edu; guweb2.gonzaga.edu

The citric acid cycle is a series of reactions that produceswoCO2, one GTP/ATPand reduced forms of FADH2 and NADH2; https://www.boundless.com ;www.narragansett.k12.ri.us
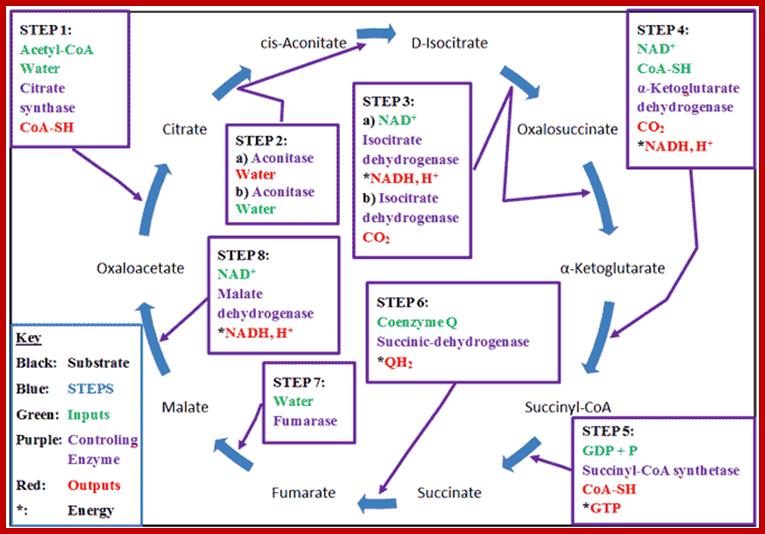
The Krebs Cycle (also known as the Citric Acid or Tricarboxylic Acid (TCA) cycle) is the process through which aerobic cellular metabolism occurs. Hans Krebs received the 1953 Nobel Prize in Medicine for his “discovery” of the citric acid cycle. This cycle involves a series of reactions involving a (1) a substrate, Oxaloacetate, that is modified in every reaction, (2) Acetyl–CoA, from which energy is extracted, (3) energy transport reactants, which collect the extracted energy, and (4) the controlling enzymes, which regulate the steps of the cycle. This cycle is ubiquitous in living organisms, single and multi-celled, both plants and animals — including humans. Organizationally, the process is often divided into 8 steps, one for each controlling enzyme, usually beginning with the combination of the Oxaloacetate substrate to the Acetyl–CoA, which is produced from either glycolysis or pyruvate oxidation. http://www.wyzant.com/
The mitochondrial matrix which fills the internal mitochondrial chambers possesses all the enzymes needed for Kreb’s cycle. The matrix also contains other enzymes required for β-oxidation of fatty acids, amino acid metabolism, protein metabolism and nucleic acid metabolism. The mitochondrial chamber is incompletely compartmentalized by the folds developed from the inner membrane called Cristae.

Inputs to Citric Acid Cycle; www.boundless.com
Then the enzyme aconitase brings about a reversible catalysis of citrate to isocitrate through an intermediate compound called aconitate by adding and removing a molecule of water
The isocitrate is then subjected to dehydrogenation and decarboxylation by an enzyme complex called isocitrate dehydrogenase to produce alfa ketogluterate.
The coenzyme involved in this reaction is mostly NAD. Once again, the activity of this enzyme is modulated by the levels of ADP/ADP ratio.
Alfa ketogluterate is a key intermediate compound in the cellular metabolism, because it is also used in the synthesis of glutamate and other amino acids. However, in citric acid cycle alfa-ketogluterate is converted to succinate in two steps reactions by an enzyme called alfa ketogluterate dehydrogenase. The catalytic action of this multiple enzyme complex is similar to that of pyruvate dehydrogenase. But the interesting reaction involved in this is the formation of high energy bond in succinyl Co.A. This high energy bond in succinyl Co.A. is used in the substrate phosphorylation of GDP to GTP.
Alpha Ketogluterate -> Succinyl Co A (energy rich bond)
Succinyl CoA + GTP +P -> Succinate + GTP
The succinate is a 4-carbon dicarboxylic acid. It is further converted to Fumerate by the action of succinate dehydrogenase, whose coenzyme FAD gets reduced to FADH2. Malonate acts as the competitive inhibitor for this enzyme. Then the fumerate is converted to malate by the addition of H2O by an enzyme called Fumerase.
Succinate +FAD-> Fumarate + FADH2
Fumerate + H2O -> Malate
Malate is then subjected to dehydrogenation by malate dehydrogenase to produce oxoloacetate.
Malate + NAD -> NADHH + OAA
The OAA thus produced, then combines with another molecule of acetyl Co. A and the citric acid cycle is repeated.
OAA + Acetyl CoA-> Citrate
At the end of Kreb’s cycle; 1 mole of glucose yields 6 C02, 10 NADH2 and 6 ATPs of which 2 ATPs are used in the first two priming steps of glycolysis.
TERMINAL OXIDATION (Oxidative Phosphorylation);
Energy rich compounds like NADH+H and FADH2 that are produced during Kreb’s cycle within mitochondria are drawn into inner cristae membrane, where they are subjected to a series of oxido reduction reactions during which energy is released. This process is referred to as terminal oxidation. As in this process ADP is phosphorylated to ATP it is also called oxidative phosphorylation. In this process the chemical energy released during electron transport is coupled for energy rich phosphate anhydride bond formation in ATP.
The process of terminal oxidation takes place in series of steps. The oxido-reductases that are responsible for this process are found in the cristae membranes as specific groups they have different red-ox potential, because of this difference, certain amount of energy is released at different oxido reduction steps. The energy released at these steps is used in the formation of high energy anhydride bonds in ATP. Such a process is termed as oxidative phosphorylation. Oxidation of one mole of NADH2 yields 3 moles of ATPs; and 1 mole of FADH2 yields 2 moles of ATPs. The unused energy is released in the form of heat.
Organization of Electron Transport Chain:
The oxido-reductive enzymes are found in the inner membrane of the mitochondria and the same are organized into 5 complexes. Complex I to complex IV are found buried within the membranes and the complex V is found on the outer surface of the oristae membrane slightly buried, but its head projects out into the mitochondrial matrix and in line with the electron transport chain.


http://oregonstate.edu/
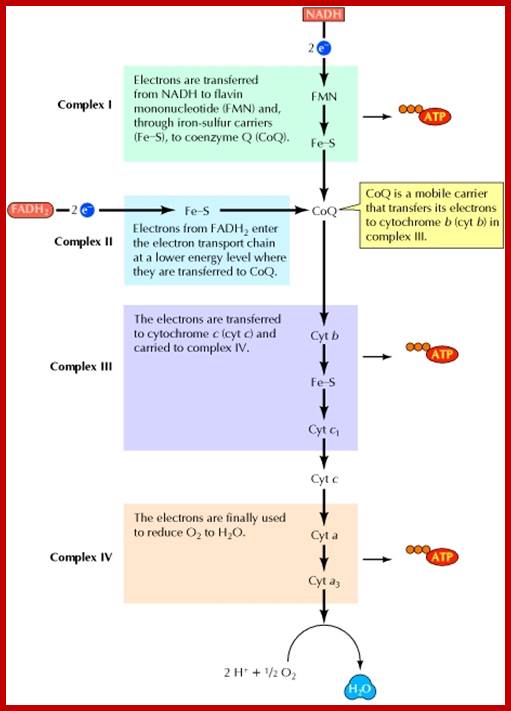
Electron transport chain: Electrons from NADH and FADH2 are transferred to O2 through a series of carriers organized into four protein complexes in the mitochondrial membrane. The free energy derived from electron transport reactions at complexes I, III, and IV is used to drive the synthesis of ATP. ; http://www.ncbi.nlm.nih.gov/

Schematic representation of the mitochondrial respiratory chain: The twelve mtDNA-encoded subunits in C. elegans are shown in red and nuclear DNA (nDNA)-encoded subunits in green. The table indicates the estimated numbers of subunits that comprise the nematode complexes. IMM, inner mitochondrial membrane; IMS, intermembrane space; Q, ubiquinone; Cyt c, cytochrome c.;www.wormbook.org

What happens when NADH binds to complex I? It binds to a prosthetic group called flavin mononucleotide (FMN), and is immediately re-oxidized to NAD. NAD is"recycled," acting as an energy shuttle. What happens to the hydrogen atom that comes off the NADH? FMN receives the hydrogen from the NADH and two electrons. It also picks up a proton from the matrix. In this reduced form, it passes the electrons to iron-sulfur clusters that are part of the complex, and forces two protons into the intermembrane space. Electron transport carriers are specific, in that each carrier accepts electrons (and associated free energy) from a specific type of preceding carrier. Electrons pass from complex I to a carrier (Coenzyme Q) embedded by itself in the membrane. From Coenzyme Q electrons are passed to a complex III which is associated with another proton translocation event. Note that the path of electrons is from Complex I to Coenzyme Q to Complex III. Complex II, the succinate dehydrogenase complex, is a separate starting point, and is not a part of the NADH pathway. Http://www.ruf.rice.edu/

ETS pathway: An inhibitor may completely block electron transport by irreversibly binding to a binding site. For example, cyanide binds cytochrome oxidase so as to prevent the binding of oxygen. Electron transport is reduced to zero. Breathe all you want - you can't use any of the oxygen you take in. Rotenone, on the other hand, binds competitively, so that a trickle of electron flow is permitted. However, the rate of electron transport is too slow for maintenance of a gradient.www.ruf.rice.edu

http://biosiva.50webs.org/
The Q/ Cycle:
- Electrons enter complex III via coenzyme QH2, which binds at a site on the positive side of the inner mitochondrial membrane, adjacent to the intermembrane space.
- QH2 gives
up one electron to the iron-sulfur center (Fe-S).
Fe-S is reoxidized by transfer of the electron to cytochrome c1, which passes it out of the complex to cytochrome c.
The loss of one electron from QH2 would generate a semi-Quinone radical, shown here as Q·, although the semi-Quinone might initially retain a proton as QH·. - A
second electron is transferred from the semi-Quinone to cytochrome bL (heme
bL), which passes it across the membrane via cytochrome bH (heme
bH) to another CoQ bound at a site on the matrix side of the
membrane.
The fully oxidized CoQ, generated as the second electron is passed to the b cytochromes, may then dissociate from its binding site adjacent to the intermembrane space. - Accompanying the two-electron oxidation of bound QH2, 2H+ are released to the intermembrane space.
In an alternative mechanism that has been proposed, the two electron transfers from QH2 to Fe-S & cyt bL may be essentially simultaneous, eliminating the semi-Quinone intermediate.
It takes 2 cycles for CoQ, bound at the site near the matrix side of the membrane, to be reduced to QH2, as it accepts 2 electrons from the b hemes and 2 H+ are extracted from the matrix compartment. In 2 cycles, 2 QH2 enter the pathway, and one is regenerated.
Overall reaction
catalyzed by complex III, including net inputs and outputs of the Q
cycle:
QH2 + 2H+(matrix side) + 2 cyt c (Fe3+) à Q
+ 4H+(outside) +
2 cyt c (Fe2+); http://biosiva.50webs.org/

http://oregonstate.edu/

http://oregonstate.edu/

http://www.bioscirep.org/

Just show the difference with electron transport in mitochondria and chloroplasts. Schematic representation of the electron transport system in the thylakoid membrane showing three possible sites of activated oxygen production.
a) Singlet oxygen may be produced from triplet chlorophyll in the light harvesting complex.
b) Superoxide and hydrogen peroxide may "leak" from the oxidizing (water-splitting) side of PSII.
c) Triplet oxygen may be reduced to superoxide by ferredoxin on the reducing side of PSI.
In Mitochondria:
Complex I consist of NADH2 dehydrogenase with FMN as the prosthetic group and 16-24 Fe-S protein.
Complex II is made up of FADH2 dehydrogenase with FAD as the coenzyme and 6-8 FE-3 containing proteins.
Complex III has heme containing cytochrome b and Cytochrome-C proteins. Cyt.C is mobile; it is also found perimitochondrial space. The release of this in to cytoplasm can lead to apoptosis. In addition, it also possesses 2 Fe-S containing proteins.
Complex IV consists of Cyt.c & Cyt.a3 oxidase proteins with 2 copper and 2 heme moieties.
Complex V is made up of F1 protein called ATP synthetase and a protein to secrete Protons. In addition, it also possesses an oligomycin sensitive conformational protein (OSCP).
Is there an evolutionary reason for the 5-electron transport complexes in plants and animals same? Is this a coincidence, or does this possibly suggest that both chloroplast and mitochondrion precursors derived their energy-obtaining mechanisms via an ancient evolutionarily-related metabolic pathway which diverged during the course of evolution?
Purple bacteria and cyanobacteria are some of the oldest known lineages (>3BY, whereas Mitochondria parallel Eukaryotic origins at about 1.4BY - and the endosymbiosis which created Chloroplasts probably came after [plants have both Chloroplasts and Mitochondria, whereas most Eukaryotes just have Mitochondria]), and since the respiratory chain and photosynthetic chain share identical proteins in cyanobacteria and purple bacteria it's not a huge leap to assume that - yup - the 5-protein structure is probably the template used by both Chloroplasts and Mitochondria because of a common source. http://biology.stackexchange.com/
Now we can calculate the end result of glucose degradation: the oxidation is coupled to a decrease of the free energy; 686 kcal/mol (= 2881 kJ/mol) are obtained by the complete oxidation of glucose. How much of this energy can the cell use?
1. Six mol ATP per mol glucose are generated (substrate chain phosphorylation). This is because all steps after the breaking down of fructose-1,6-phosphate have to be counted twice (once for each of the two resulting C3 molecules), so it is 3 x 2 ATPs. Of these six ATPs, two are needed to start glycolysis. That leaves four.
2. During the course of glycolysis up to acetyl-CoA, 2 x 2 NADH + H+ are generated. An additional 3 x 2 NADH + H+ and 1 x 2 FADH2 are produced in the citric acid cycle. One NADH + H+ gives three, one FADH2 two ATPs when fed into the respiratory chain. This sum up to 34 ATPs plus the 4 ATPs of glycolysis. A total of 38 molecules of ATP are thus gained by the cell's degradation of one molecule of glucose. Since each energy-rich bond of ATP contains 7.3 kcal/mol (= -30.6 kJ/mol), the 38 ATP equal 277 kcal/mol (ca 1163 kJ/mol). This is 40.6% of the theoretically possible gain. The other 59.4 percent are set free as heat. This is a very high percentage compared to the gain of technical machines like steam or petrol engines that is around or below 20 percent. https://s10.lite.msu.edu

www.antranik.org

Decrease of
free energy (delta G)
during glycolysis and respiration.
Conversion factors: 200 kcal = 1680 kJ ; 600
kcal = 2520 kJ.
;https://s10.lite.msu.edu

Energy released at each of the transfer steps; http://dwb4.unl.edu/


Oxidative phosphorylation; http://english.eagetutor.com/
Complex-I; Referred as NADH dehydrogenase
Electron donor is NADH
Electron acceptor is ubiquinone (Q) and converts
to ubiquinol (QH2); Complex
II; Referred as Succinate dehydrogenase
complex Electron acceptor is ubiquinone (Q) and converts to
ubiquinol (QH2) Electron donors are fatty acids, glycerol 3-phosphate, Complex II
Referred as Cytochrome bc1 complex Electron Donor is
Ubiquinol (QH2) Electron acceptor is cytochrome C, Complex IV
Referred as Cytochrome C Oxidase Electron donor is Cytochrome
C Electron acceptor is molecular oxygen. NADH → Complex I → Q → Complex III→ Cytochrome C → Complex IV → O2;
The electron transport chains of both the light reactions of photosynthesis (in plants) and oxidative phosphorylation (in animals) both contain 5 complexes including ATP synthase, as shown below.


http://dwb4.unl.edu/;https://s10.lite.msu.edu
Oxidative Phosphorylation:

Mitochondria: http://biology.stackexchange.com/https://s10.lite.msu.edu

Comparison of electrons flow in mitochondria and chloroplasts; Photosynthesis; Link between PSII and PSI; https://s10.lite.msu.edu
The first four complexes from complex I to complex IV are vectorially disposed within the core of inner mitochondrial membrane, but the complex V is located on the surface of the inner membrane in such a way, the head i.e. F1 is projected out into the mitochondrial matrix and the basal part is buried in the lipid core of the membrane. But the OSCP acts as a stalk between Fo and F1 part of the ATP synthase enzyme complex.

ATP synthase complex; www.imgarcade.com
Such particles are uniformly distributed over the surface of the inner membrane. However, this complex when it is free from the membrane (probably only) with the head portion, it acts as the hydrolysing ATPase enzyme. On the contrary, if it is associated with the membrane (with fo & OSCP) it functions as ATP synthetase or ATP synthase. Along with the above said complex they are some other components in the membrane which are free and labile. In fact, they act as intermediary components between complexes. They are coenzyme Q and cyt. C; the coenzyme Q acts as a shuttle between complex I and complex III and complex II and complex II, but not between complex I and II, similarly, cyt. C acts as a shuttle between complex III and complex IV. During terminal oxidation process, NADH2 reacts with complex I but not with any other complex. Similarly, FADH2 reacts with complex II only.
MECHANISM OF TERMINAL OXIDATIONS
When NADH2 reacts with complex I, NADH2 gets oxidized to NAD and the FMN of complex I gets reduced to FMNH2. Because of the difference in oxido-reduction potential between the oxidant and the reductant ̴2.2 K.cals/mole of energy is liberated and the same is used in the formation of ATP by oxidative phosphorylation process. On the other hand, FADH2 reacts with complex II where the prosthetic group of complex II enzyme i.e. FAD gets reduced to FADH2. The redox potential between these two is almost equal and the free energy released is nil.
The coenzyme Q which acts as the shuttle between complex I and complex III and complex II and complex III gets reduced to QH2. The coenzyme Q accepts hydrogen either from FMNH2 or FADH2. The energy released in this redox reaction is negligible to produce any energy rich bond. The most interesting step is the reaction between QH2 and complex III, where the reaction results in the separation of H protons and electrons. Only the electrons are channeled to cyt. B of the complex III and H ions is released into the membrane. As cyt. B Fe3 + accepts the electron and it gets reduced to Cyt.B fe2+. In this redox reaction, some amount of energy is released but not sufficient to produce an ATP. The reduced cyt. B Fe 2+ reacts with oxidized cyt.c1 FE3+. Where cyt.bfe2 gets oxidized and cyt.c Fe3 gets reduced to cyt.Gfe2. The energy released in this redox couple is about 9.9 K.cals/mole and the same is used up n the formation of another energy rich ATP molecule.
Then cyt.c1fe2+ reacts with cyt.C fe3+, where cyt.cfe3 gets reduced to cyt.C fe2+ and cyt.c1fe2 gets oxidized to cyt.c1fe3+. The free energy released in this reaction is not sufficient to produce ATPs. From cyt.cfe2+ the electrons are transported through cyt.a and cyt.a3. It is during the redox reactions between cyt. A and cyt.a3, hereby 23.8 K.cals/mole of energy is released and the same is utilized in the generation of ATP. Finally, the electrons from cyt.a3fe2+ are transferred over to o2, which simultaneously receives 2H+ to produce a water molecule.
It is very important to remember that during electron transportation, only one electron is shuttled between the redox couples at a time, but the flow of electrons is very fast. However, the synthesis of ATPs is calculated for a pair of electrons flow at any given step. Thus the oxidation of NADH+H yields 3 ATPS and FADH2 yields 2 ATP.
The presence of 3 sites in the electron transport chain at which ATPs generate has been identified by using drugs such as rotenone, antimycin and cyanide. Based on these studies, it has been identified that rotenone binds to complex I, antimycin binds to complex III and cyanide binds to cytochrome oxidase complex and prevents the electron flow and also prevents ATP formation. So the sites of ATP formation are between NADH2 and FMN of complex I, Cyt.b and Cyt.c1 of complex III and Cyt.a and Cyt.a3 of the complex IV.
All in all, one mole of glucose on complete glycolysis, Kreb’s cycle and terminal oxidation yields a total of 40 ATPs of which 2 ATPs are used in the early glycolytic reactions. So the net gain in terms of ATP production is 38 per mole of glucose. In actuality 1 mole of glucose possesses 686 K.cals energy out of which 38x 7.2 K.cals of energy is trapped is in the formation of 38 ATPS. The rest of the energy is released as heat. Thus the efficiency of aerobic type of biological oxidation in terms of energy conserved is hardly 39.3%, it is not 100%, but energy released as heat is required to maintain the body temperature. Animals and plant that live in Arctic and Antarctic regions do store lot of fat, during hibernation period they use that fat in oxidative process, where the released energy in electron transport is used. It also uses electron transport process only to release heat rather ATPs.
Mechanism of ATP Synthesis:
Though the energy is released at different sites during the electron transport the mechanism by which the energy is utilized is in the formation of energy rich phosphate bond in ATP is not completely elucidated. In this text book, three important theories have been discussed.
HIGH ENERGY CHEMICAL INTERMEDIATE THEORY
The protagonists of this theory namely Slater, Britten chance and others believe that the energy released at specific sites in the electron transport chain is trapped as an high energy bond in an unknown compound called X. Then the same energy rich bond is transferred to inorganic phosphate, which is then transferred to Pi and than to ADP to generate ATP. This concept was invoked, because the formation of high energy bond can be inhibited by the addition of an uncoupler of oxidative phosphorylation called dinitrophenol (DNP), where all the energy released during electron transport is lost in the form heat, interestingly in this process the flow of elctron is not inhibited but ATP synthesis is completely prevented. This indicates the presence of some intermediate compound or component which acts as the receiver of energy for coupling reactions in generating ATPs. Unfortunately, the presence of such energy rich substance has not been detected so far. Hence, this theory is not favored by many scholars in this field.
CHEMIOSMOTIC THEORY
Peter Mitchell proposed this theory and it was supported by many. But there are others who vehemently argue against this theory. In spite of it, he was awarded a Nobel Prize in 1978, for his significant contribution in the field of bioenergetics; no other field has generated so much controversy and heated debates as in the case of mitochondrial bioenergetics.


A model of the way protons are translocated through the inner mitochondrial membrane. Please notice that the protons are not passed on but that OH-ions are set free at the outside of the membrane while protons are given away into the matrix at the inner surface. This results in a seemingly directed movement of the protons. The ATP synthase (also known as ATPase) has two neighbouring binding sites for ADP and phosphate. http://dwb4.unl.edu/

http://dwb4.unl.edu/;https://s10.lite.msu.edu
In 1961 postulated P. MITCHELL that protons are translocated through the membrane in a directed way. This results in a pH gradient.
The passage back occurs via a specific proton channel. This passage is coupled to ATP-synthesis, using the potential energy of the proton gradient for the formation of the third phosphate bond of ATP.
We can now calculate the end result of glucose degradation: the oxidation is coupled to a decrease of the free energy; 686 kcal/mol (= 2881 kJ/mol) are obtained by the complete oxidation of glucose. How much of this energy can the cell use?
1. Per mol glucose are six mol ATP generated (substrate chain phosphorylation). This is because all steps after the breaking down of fructose-1,6-phosphate have to be counted twice (once for each of the two resulting C3 molecules), so it is 3 x 2 ATPs. Of these six ATPs are two needed for to start glycolysis. That leaves four.
2. During the course of glycolysis up to acetyl-CoA are 2 x 2 NADH + H+ generated. An additional 3 x 2 NADH + H+ and 1 x 2 FADH2 are produced in the citric acid cycle. One NADH + H+ gives three, one FADH2 two ATPs when fed into the respiratory chain. This sums up to 34 ATPs plus the 4 ATPs of glycolysis. A total of 38 mol ATP are thus gained by the cell's degradation of one mol glucose. Since each energy-rich bond of ATP contains 7.3 kcal/mol (= -30.6 kJ/mol) do the38 ATP equal 277 kcal/mol (ca 1163 kJ/mol). This is 40.6% of the theoretically possible gain. The other 59.4 percent are set free as heat. This is a very high percentage compared to the gain of technical machines like steam or petrol engines that is around or below 20 percent. http://dwb4.unl.edu/
Mitchell’s chemiosmotic hypothesis is based on the assumption that NADH2 and FADH2 are drawn into inner mitochondrial membrane by the electron transport system and then the protons (H+) are released at the outer surface of the membrane into periplasmic space and the electrons are transported along the electron transport chain. Though the electrons, that are transported along the chain end up in oxygen, the extrusion of positively charged H+ at the outer surface of the inner mitochondrial membrane, builds up greater positive charge outside and negative charge inside the membrane. The membrane potential thus generated provides a strong proton motive force for the generation of high energy bonds between ADP and Pi, which results in the formation of ATP. The differential charge is believed to be neutralized by the synthesis of ATP, during which the OH ad H produced are released to outside and inside of the time of anhydride bond formation between ADP and Pi.
One of the best evidence that has been presented in favor of this hypothesis is from Jogendorf’s experiments on chloroplasts. After soaking the isolated chloroplasts to saturation in a buffered medium with pH 4 (acid) in dark, when the said chloroplasts are transferred to another buffered medium with alkaline pH 8.5 containing ADP and Pi, resulted in a burst of ATP synthesis. This explains how membrane potential with high negative charges and positive charges operating on the opposite surfaces of the membranes can generate energy rich bond between ADP and Pi to synthesize ATPs.
In spite of overwhelming circumstantial evidences, research workers who were contradicting the chemiosmotic hypothesis developed new and sophisticated microelectrodes to measure the smallest change in the pH in the shortest possible time. With such new tools and techniques they measured the time, required for the generation of proton motive forces across the mitochondrial membranes and the time required for the synthesis of ATP in mitochondria. Such studies demonstrated that the time required for the building up of proton motive force to generate ATB molecules runs to a few milliseconds, but in actuality ATPs are synthesized within a few nano seconds after the elctron transport. The time lag between the generation of sufficient proton motive force across the mitochondrial membranes to generate ATP synthesis, the actual time required for ATP synthesis due to electron transport within the membrane, is found to be a strong evidence against chemiosmotic hypothesis. In similar experiments people have shown that blocking proton transfer does not prevent the synthesis of ATP into mitochondrial inner membranes. Furthermore, in plant cells, where auxin causes secretion of H ions outside the plasma membrane to such an extent the pH at the outer surface of the membrane falls from 6.8 to 4.0. Though ADP and Pi are available at the site ATPs are not synthesized some of there objections are solid and chemiosmots hypothesis has no answer to them.
CONFORMATIONAL CHEMICAL COUPLING THEORY
In actuality, the conformational hypothesis was initiated by David Green et al. But later Boyer and others developed a new hypothesis and gave new dimensions to the above said theory. In Mitochondria, in its resting state when internal membranes are in a relaxed condition, but when mitochondria are fully charged with ATP, the internal cristae membranes show maximum contraction where even ATP synthetase complex with its head and stalk show drastic changes in their morphological conformations. The analysis of internal membrane protein components of mitochondria indicates the presence of some contractile proteins similar to actions. Furthermore, such proteins are believed to act as semiconductors of electronic energy.
This hypothesis envisages that during the electron flow from high energy state to lower levels, because of the difference in redox potential between the oxidants and reductants of different complex, certain amount of energy is released at different sites. The same energy is low drawn into some contractile proteins associated with ATP syntheses. By virtue of energitization of these proteins, the ATP synthetase undergoes conformational change. As ADP and Pi are already bound to specific sites on the surface of ATP synthetase, the conformational change brings the ADP and Pi together and the energy found in contractile proteins is used up in the formation of energy found in contractile proteins is used up in the formation of energy rich bonds between ADP and Pi. Once the energy is used up the contractile proteins and ATP syntheses enzymes relax to their almost simultaneously original state, ATP is released from the enzyme surface. And this process is repeated again and again to produce more ATP molecules. This process takes place very rapidly. Hence this hypothesis appears to explain all the observed facts about the mechanism of ATP synthesis. The explanation is very simplistic and seminal. Recently the mitochondrial people consider the proton motive force is very important in producing ATP molecules. Accordingly, they consider that the proton binding to ATP synthase, particularly binding and release of them determines the rate of ATP synthesis.

This diagram of ATP Synthase within the mitochondrial inner membrane shows two major strutural components Fo and F1 and the subunits within each. Permission for use of this image is being sought at <http://pathology.mc.duke.edu/research/postpizzo.html>, where it can be found in its original context. http://www.bio.davidson.edu/
ATP Synthase- A vital protein complex used in oxidative phosphorylation to generate ATP;
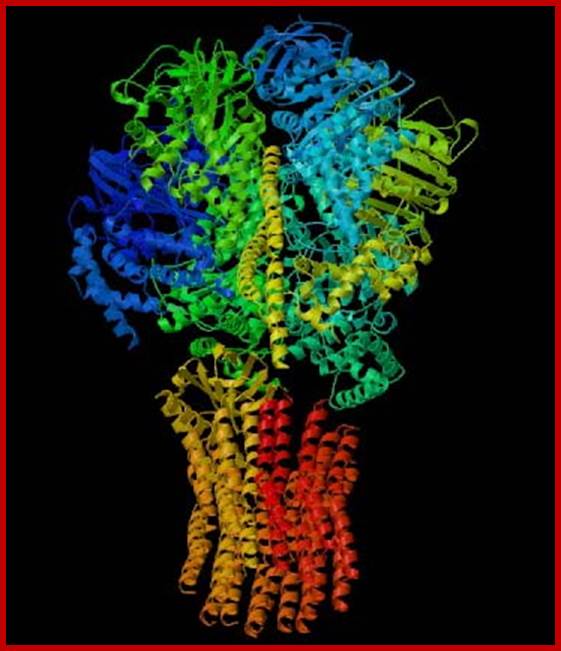
This 3-D image of ATP Synthase illustrates the protein's complex tertiary structure. http://www.bio.davidson.edu/
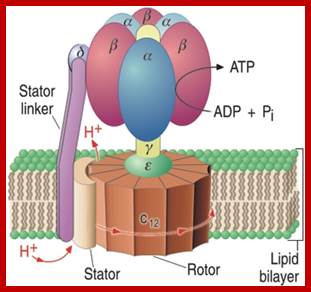
ATP synthase, known as Vth complex in oxidative phosphorylation; is located in the inner mitochondrial membrane. It is associated with electron transport chain, this creates proton (H+) concentration gradient across the membrane; which acts as a proton motive force (according to Peter Mitchell), which drives ATP synthesis as protons flow across the membrane. Among the four types of ATP synthases, the AT synthase found in mitochondria contain a peripheral domain called F1 and integral domain called Fo; which pump protons only in one direction.
ATP synthase complex consists of total 20 protein subunits. The F1 has nine subunits of five different types; 3 beta and 3 alpha alternating to each other for a total of six subunits. The knoblike portion of the F1 is a flattened sphere of 8nm/10nm. A central shaft runs through the center of these six subunits. The delta and epsilon subunits attached to gamma, thus help in linking F1 domain to the Fo domain. The central stalk rotates 50-100 time per second.
The amino acid sequences of all three beta subunits are identical, but their conformations differ, mainly because of the association of the gamma subunit with just one of the three. The ATP-ADP binding sites of the three beta subunits differ and are labelled beta-ATP, beta-ADP, and beta-empty. This difference in binding is critical to the protein's mechanism. For every three protons that flow through ATP Synthase, the complex rotates one position (due to the rotation of the gamma subunit) and one ATP molecule is formed. The three beta subunits interact in such a way that when one subunit releases ATP and become beta-empty, its neighbor to one side must assume the beta-ADP conformation and the neighbor one the other side must assume the beta-ATP conformation. Thus, for each complete rotation, three ATP are synthesized and released (Nelson et al., 2000). The image below shows this rotation.


http://www.mikeblaber.org/
This image shows the conformations of the beta subunits as the complex rotates. For every 3 protons that go through the enzyme, one ATP is produced and released. This image was found at the same site as Figure 4 and permission is currently being sought.
As mentioned earlier there are four classes of ATPases and ATP synthases, they are F-type, they vary slightly from one another. Furthermore, V-Type and F-Type ATPases in particular share common structural components. Like ATP Synthase, V-type ATPases have a peripheral domain (V1) and an integral membrane component (Vo). V1 has seven different subunits, with three A and three B around a central stalk D. Vo also has three types of subunits with multiple copies of subunit c. Obviously this structure is very similar to that of F-type ATPases, but V-type ATPases are located in vascular membranes of higher plants and fungi and in lysosomal, endosomal, and secretory vesicle membranes of animals rather than the mitochondrial inner membrane (Nelson et al, 2000).
* Creation of the ATP SYNTHASE Motor *
The three a/b complexes are identical to each other, but are able to exist in three different conformational states:
- The "open" conformation (conformation O). Low affinity for ATP, ADP or phosphate ion, and is catalytically inactive
- The "loose" conformation (conformation L). Binds ATP, ADP and phosphate ion loosely; no catalytic activity
- The "tight" conformation (conformation T). Binds ATP, ADP and phosphate tightly, is catalytically active
The conformational states of the three a/b complexes in the "stator" are influenced by asymmetric interactions with the "rotor" and sequentially cycle through these states:


http://www.sigmaaldrich.com/;http://biosiva.50webs.org/; http://www.rcsb.org/; www.life.illinois.edu

Mr~500,000 and ~8.5nm Sphere attached to matrix side of inner membrane; FoF1 ATPase; http://www.bmb.leeds.ac.uk/
The complete assembly contains at least 12 different types of polypeptide chain, several of which are present in multiple copies. The catalytic head group is connected by an oligomycin sensitive stalk to a proton conducting baseplate in the mitochondrial inner membrane. Three protons are thought to pass through the membrane from the external P phase to the internal N phase for each molecule of ATP manufactured by the complex.
The F1 head group contains three nucleotide binding sites, and the enzyme probably performs a three-phase catalytic cycle. In the first phase, ADP and phosphate bind to one active centre, which catalyzes the formation of bound ATP. This step is energetically possible because the free energy released by tightly binding the ATP to the active centre compensates for the instability of the new phospho anhydride bond. The energy from the proton motive force is required to prise the ATP from the active centre.
The F0 base piece embedded in the mitochondrial inner membrane is a molecular turbine driven by the trans-membrane proton gradient. Proton entry forces a central camshaft to rotate within the F0 baseplate and the F1 head group, altering the subunit conformation as this movement takes place. A second, off-center protein tether connects the head group to the base piece and prevents the head piece spinning uselessly as the central shaft rotates. Energy is transmitted to the catalytic subunits in the ATP synthase F1 headpiece by the rotation of the camshaft. The "cam" distorts the protein subunits, destroying their ability to bind ATP. The energy input is used to drive ATP release, not for bond formation. http://www.bmb.leeds.ac.uk/


Exploring the F0 Structure; David Goodsell; Cars run on gasoline, bio-life runs on ATP; http://www.sigmaaldrich.com/;http://biosiva.50webs.org/; http://www.rcsb.org/; www.life.illinois.edu

Structure scheme of F1ATPase; The cut through the hollow F1ATPase head shows three pairs of subunits (alpha, beta heterodimers) and the rotating gamma subunit in the center. Each alpha, beta-pair carries one catalytic center (in the cleft) and an additional buried non catalytic nucleotide center (orange). Only one subunit pair is associated with the delta-subunit (epsilon in Micrococcus luteus), which is part of the proposed "second stalk" structure; http://mpsd.de/ATP-Synthase.html

Spinning of the gamma subunit is caused by movement of protons through the c ring of the F0. Tight links between the c ring and the gamma and epsilon subunits cause them to rotate as well. Each 360-degree rotation of the c ring generates 3 ATPs. http://oregonstate.edu/


Proton transport facilitated by basal Fo-C complex and rotation of central rod Gamma rotor; http://oregonstate.edu/

{kind=link}
Spinning of the gamma subunit is caused by movement of protons through the c ring of the F0. Tight links between the c ring and the gamma and epsilon subunits cause them to rotate as well. Each 360-degree rotation of the c ring generates 3 ATPs. http://oregonstate.edu/
The structure includes the F0 electric motor. In this picture, we are looking down the axis of rotation, as if we, where looking down at the top of the picture on the first page. The rotor is composed of 12 identical protein chains, colored blue here, and the ion pump is a single chain, colored red. The pump has an arginine amino acid that hands off a hydrogen ion to aspartates on the rotor. Aspartate amino acids typically have a negative charge, but since the rotor is surrounded by membrane lipids, this would be very unfavorable. So, the rotor only turns when the aspartates have hydrogen attached, neutralizing their charge. Hydrogen ions take a convoluted path through the F0 motor, turning the rotor in the process. They are gathered by a chain of amino acids in the pump, and transferred to the arginine. The arginine passes the hydrogen to the rotor, which turns all the way around. Then the hydrogen is offloaded by other amino acids on the pump, and finally passed to the opposite side of the membrane. The exact path of the hydrogen ions through the pump is still a matter of intense study. David Goodsell
- rotation of the central shaft within F1 to coordinate the actions of three cooperative, alternating catalytic nucleotide sites, and
- rotation of subunits within F0 during energy-driven proton transport.
Remarkably, energy coupling by the ATP synthase is very efficient and reversible: respiration-driven proton transport through F0 can drive net synthesis of ATP, but net hydrolysis of ATP can also be used to drive proton transport in the opposite direction, thereby generating a transmembrane, electrochemical proton gradient.
ANAEROBIC RESPIRATION
Respiration without utilizing oxygen or in the absence of oxygen is called anaerobic respiration. Yeasts and some bacteria respire even in the absence of oxygen and ethanol or lactate is produced as the end products. Surprisingly, even higher animals exhibit anaerobic respiration under certain conditions where oxygen supply is inadequate.

www2.estrellamountain.edu

Yeast cells are capable of performing respiratory processes both aerobically and anaerobically. Under aerobic conditions, yeast cells possess 150-200 active mitochondria per cell. On the other hand, if such are subjected to anaerobic conditions, the number of mitochondria reduces dramatically to 1 to 2 per cell. But with the restoration of aerobic conditions, the original number of mitochondria will be restored. Thus yeast cells show remarkable adaptations.

Glycolysis-Chemwiki; http://chemwiki.ucdavis.edu/



www.shmoop.com; www.cheesescience.com
During anaerobic respiration, glucose is subjected to same biochemical reaction as in the case of glycolytic steps. For that matter there is no difference between anaerobic and aerobic glycolytic pathways. The pyruvates produced as the end products of glycolytic reactions, are first decarboxylated and then reduced to ethanol by alcohol dehydrogenase enzymes. In this process, I mole of glucose yields 2 moles of CO2, 2 moles of ethanol and the net gain in ATP is 2 moles. This process is also called partial biological oxidation because a part of the glucose is lost as CO2.
In the case of lactate bacteria, the pyruvate produced at the end of glycolysis is converted to lactate by the action of lactate dehydrogenase activity. Here, even CO2 is not released. Hence this kind of respiration is often referred to as intramolecular respiration. The net gain of ATP in this process is also 2.
Even in higher mammals, muscular respiration under inadequate supply of oxygen during intense muscular work, pyruvate is converted to lactate. The accumulation of lactate in the muscles causes fatigue and pain.
During respiration it is not just carbohydrates, especially glucose is not the sole source, but fatty acids, another important source for energy; there are many other organic molecules are drawn and used for generation ATP and other similar energy.
Oxidation of Fatty Acids:
Fats (triacylglycerols); Fatty acids are the major storage form of lipids. The first step in their utilization is their breakdown to glycerol and free fatty acids. Each fatty acid is joined to coenzyme A, yielding a fatty acyl-CoA at the cost of one molecule of ATP (Figure 2.36). The fatty acids are then degraded in a stepwise oxidative process, two carbons at a time, yielding acetyl CoA plus a fatty acyl-CoA shorter by one two-carbon unit. Each round of oxidation also yields one molecule of NADH and one of FADH2. The acetyl CoA then enters the citric acid cycle, and degradation of the remainder of the fatty acid continues in the same manner. http://www.ncbi.nlm.nih.gov/
Energy in the form of ATP can be derived from the breakdown of other organic molecules, with the pathways involved in glucose degradation again playing a central role. Nucleotides, for example, can be broken down to sugars, which then enter the glycolytic pathway, and amino acids are degraded via the citric acid cycle. The two principal storage forms of energy within cells, polysaccharides and lipids, can also be broken down to produce ATP. Polysaccharides are broken down into free sugars, which are then metabolized as discussed in the previous section. Lipids, however, are an even more efficient energy storage molecule. Because lipids are more reduced than carbohydrates, consisting primarily of hydrocarbon chains, their oxidation yields substantially more energy per weight of starting material.
Fats (triacylglycerols) are the major storage form of lipids. The first step in their utilization is their breakdown to glycerol and free fatty acids. Each fatty acid is joined to coenzyme A, yielding a fatty acyl-CoA at the cost of one molecule of ATP. The fatty acids are then degraded in a stepwise oxidative process, two carbons at a time, yielding acetyl CoA plus a fatty acyl-CoA shorter by one two-carbon unit. Each round of oxidation also yields one molecule of NADH and one of FADH2. The acetyl CoA then enters the citric acid cycle, and degradation of the remainder of the fatty acid continues in the same manner. http://www.ncbi.nlm.nih.gov/

http://www.ncbi.nlm.nih.gov/books
Oxidation of fatty acids. The fatty acid (e.g., the 16-carbon saturated fatty acid palmitate) is initially joined to coenzyme A at the cost of one molecule of ATP. Oxidation of the fatty acid then proceeds by stepwise removal of two-carbon units as acetyl molecules; The breakdown of a 16-carbon fatty acid thus yields seven molecules of NADH, seven of FADH2, and eight of acetyl CoA. In terms of ATP generation, this yield corresponds to 21 molecules of ATP derived from NADH (3 × 7), 14 ATPs from FADH2 (2 × 7), and 96 from acetyl CoA (8 × 12). Since one ATP was used to start the process, the net gain is 130 ATPs per molecule of a 16-carbon fatty acid. Compare this yield with the net gain of 38 ATPs per molecule of glucose. Since the molecular weight of a saturated 16-carbon fatty acid is 256 and that of glucose is 180, the yield of ATP is approximately 2.5 times greater per gram of the fatty acid—hence the advantage of lipids over polysaccharides as energy storage molecules; http://www.ncbi.nlm.nih.gov/books
MEASUREMENT OF RESPIRATION
Respiration can be measured quantitatively or qualitatively in terms of either the uptake of oxygen or the liberation of C2 with reference to the substrate used in a given time. Such studies can be made by using whole cells or parts of the plant body like leaves, floral buds, embryos, etc. It is also possible to isolate mitochondria from any part or parts of the plant body and the uptake of O2, release of CO2 or the P/O2 ratio can be determined.
Instruments like oxygen polarograph, Warburg’s manometers can be used to determine the oxygen uptake, O2 output or P/o2 ratio under various parameters.
RESPIRATORY QUOTIENT RQ
The ratio between the amount of CO2 evolved and the amount of oxygen consumed by a given weight of tissue in a given time at standard temperature is called respiratory quotient (RQ)
RQ CO2 released / O2 consumed
Determination of RQ of a tissue gives valuable information about the rate of respiration and the substrate used. By just measuring RQ it is possible to find out what substrate is consumed during respiration. Generally, carbohydrate gives RQ value to be 1, fats 0.7, proteins 0.8 to 0.99 and organic acids like malate, tartarate oxalate, etc., more than 1.
Carbohydrates: Glucose: C6H12O6 + 602 -> 6C02 + 6HO2
FACTORS THAT CONTROL RESPIRATION:
Temperature: most of the biochemical processes mediated by enzymes or not are dependent on the temperature of the media. This is because temperature provides energy for the movement of molecules within the medium which in turn has an important effect on the frequency of collision between reactants and the enzyme. Thus the rate of the reaction is controlled by the prevailing temperature. Majority of the plants exhibit 25-30 degree as the optimal temperature for their activity but some plants living near hot springs show 40°C-60°C degree as the optimal temperature. Higher temperature affects the biochemical process because enzymes get denatured and loose their functional abilities.
OXYGEN
The availability of oxygen ultimately determines the respiratory process which may be aerobic or anaerobic type. The concentration of oxygen in the surrounding medium has a profound effect on the rate of respiration because oxygen acts as the oxidant, where it receives electrons during terminal oxidation. Increase and decrease in the concentration results in the increase and decrease I the rate of respiration. However, some cells respire both under aerobic and anaerobic conditions. But in certain cases like yeast cells, if the cells that are respiring under aerobic conditions, transferred to anaerobic media, the process of respiration immediately stops. Interestingly, the number of mitochondria in the yeast cells goes down from 200 to 1 or 2 per cell. But when such cells are transferred to aerobic condition, within the next 30 minutes or so, mitochondria regenerate and multiply and restore the normal process of respiration. On the contrary, if the cells that are growing under strict anaerobic condition are suddenly transferred to aerobic condition the rate of respiration goes down instead of increase in the rate. This effect is called Pasteur’s effect. This peculiar behavior has been explained as due to the effect of oxygen on glycolytic pathway. This is a transitory effect, with time the normal rate of respiration is restored. However, in leaves, increase in oxygen initiates photo respiration.
GLUCOSE REPRESSION
Similar to oxygen’s effect, in yeast cells, higher concentration of glucose brings about the repression of respiratory process; such a process is called glucose repression. During glucose repression, the mitochondria are degraded by the lysosomal activities. Glucose repression persists for a period of 60-90 minutes, and then mitochondria regenerate and restore the normal process of respiration. This effect is virtually similar to that of catabolitic repression fund in bacteria.
CARBON DIOXIDE
In animals higher concentration of CO2 has deleterious effect, because it brings about a change in the pH in the body fluid. In plants, however, CO2 has no direct effects. In daytimes, most of the CO2 available is used in photosynthesis, but at nights the concentration of CO2 increases. As a result of it, stomata close, thereby it affects the availability of oxygen to plants but it does not impose any ill effects on the plants. Nonetheless, CO2 brings about an inhibitory effect on ethylene induced cyanide insensitive respiration.
PHYTOHORMONES:
Plant hormones such as IAA and GA generally enhance the rate of respiration due to certain changes in membrane permeability and also due to the activation of some respiratory enzymes. At later stage, as phytohormones activate and enhance transcription and translation, the rate of respiration also increases because of the grater demand for the energy. As most of the growth promoting plant hormones accelerate cellular metabolism leading to growth, the rate of respiration also increases. On the other hand, hormones like ethylene, which induces fruit ripening, enhances the cyanide insensitive respiration to climactric state. This brings about many metabolic changes leading to softening and sweetening of the fruits. Even wounding of plant parts induces and enhances the rate of respiration because of the activity of traumatic acid. Due to the action of traumatic acid, the surface cells where the wound is inflicted, become meristematic and start cell divisions. So the cells require more energy for cellular metabolism and multiplication. That is why the rate of respiration increases at region of wound. However, growth inhibiting hormones like abscisic acid (ABA) lowers respiratory activity is plant cells.
PLANT ORGANS
The whole plant cannot be taken as a unit for measuring or determining respiratory activity. Unicellular plants can be used for such measurements. But higher plants with complex organizations exhibit different rates of respiration activity at different stages of growth and development. While germinating seeds, young leaves, apical buds, root tips, floral buds, developing inflorescence and such others exhibit higher rate of respiration, dormant seeds, dormant buds, mature fruits and seeds show minimum respiratory activity.
OXIDATIVE PENTOSE PHOSPHATE PATHWAY
This pathway is called by other names like phosphogluconate pathway or hexose monophosphate shunt. B.L. Horeker and E. Racker have made extensive studied on this pathway. Both plant and animal cells exhibit this pathway in cytoplasm outside the mitochondria. However its activity depends upon the plants requirements. In cellular metabolism, pentose sugars are required for the synthesis of nucleotides; NADPH2s are required for various reductive processes like fatty acid synthesis, reductive animation, nitrate or nitrite reduction etc. Under such demanding conditions, hexose monophosphate shunt is activated and it provides the required intermediary compounds. Thus this pathway is found to be very useful in providing many intermediary products and reducing power required for other metabolic processes.
6 Glucose 6-P+12 NADP+7H2) – 5 glucose 6-P+6CO2+12 NADPH2+Pi
For every 6 moles of glucose phosphates used, 6 moles of CO2, 12 moles of NADPH2 and one mole of phosphates are released, thereby 1 mole of glucose is completely oxidized to 6 moles of CO2 but 5 moles of glucose are recovered. Hence, equation can be rewritten as
Glucose 6-P + 12 NADP + 7H2O –> 6 CO2 + 12 NADPH2+Pi
To begin with, glucose 6-P is converted to phosphogluconlactone by the activity of glucose 6, P dehydrogenase enzyme which is called as Zwischenferment or Horecker Racker enzyme. In this process, one NADHP2 is produced.
The phosphogluconate, thus produced, is immediately hydrated to produce 6phosphogluconate. Then this is subjected to successive dehydrogenation and decarboxylation steps to produce ribulose 5, Ps by the enzyme activity of phosphogluconate dehydrogenase enzymes. Then ribulose 5P is converted to ribose 5, P by isomerase enzymes. Once ribose and ribulose are produced, they are subjected to cyclic reactions.
The following is the diagrams presented by various authors for you to appreciate and your comments.
